Department of Obstetrics, Gynecology, and Reproductive Sciences
University of Texas Health Science Center at Houston
Houston, Texas
The same gene also has alleles that cause an elevated-metabolizer phenotype antibiotic yeast purchase myambutol 600mg with amex, which has been correlated with increased susceptibility to cancer virus zapping robot best myambutol 800 mg. Clearly antimicrobial over the counter purchase 400 mg myambutol with amex, before selecting patients for clinical trials of new drugs infection rate of ebola discount myambutol 400mg line, it would make sense to screen candidates for their drugmetabolizing phenotype and most large pharmaceutical companies now do this antibiotic resistance report 2015 discount 600mg myambutol mastercard. As is well known antibiotics for dogs abscess 400 mg myambutol for sale, certain microbes quickly develop resistance to the antibiotics in current use and this means that new antibiotics are required. The traditional way of obtaining new antibiotics is the screening of new microbial isolates from nature. Another way is to identify new cellular targets and screen chemical libraries for inhibitory activities. A number of methods for identifying key genes involved in pathogenesis have been developed and these are described later in this section. In contrast to bacteria and fungi, viruses are not sensitive to antibiotics and few therapies have been available. However, this could change with the development of antisense drugs, as described in the previous section (p. Where suitable therapies do exist, it can be advantageous to know the identity of the pathogen as soon as possible. For many pathogens, prevention is much better than cure, and hence vaccines are of great value. Genemanipulation techniques have greatly facilitated the development of new vaccines, as described below. Novel routes to vaccines An effective vaccine generates humoral and/or cellmediated immunity, which prevents the development of disease upon exposure to the corresponding pathogen. This is accomplished by presenting pertinent antigenic determinants to the immune system in a fashion which mimics that in natural infections. Conventional viral vaccines consist of inactivated, virulent strains or live, attenuated strains, but they are not without their problems. For example, many viruses have not been adapted to grow to high titre in tissue culture. There is a danger of vaccine-related disease when using inactivated virus, since replication-competent virus may remain in the inoculum. Finally, attenuated virus strains have the potential to revert to a virulent phenotype upon replication in the vaccinee. This occurs about once or twice in every million people who receive live polio vaccine. Given the ease with which heterologous genes can be expressed in various prokaryotic and eukaryotic systems, it is not difficult to produce large quantities of purified immunogenic material for use as a subunit vaccine. A whole series of immunologically pertinent genes have been cloned and expressed but, in general, the results have been disappointing. Perhaps it is not too surprising that subunit vaccines produced in this way do not generate the desired immune response, for they lack authenticity. Being inert, they do not multiply in the vaccinee and so they do not generate the effective cellular immune response essential for the recovery from infectious disease. Recombinant bacterial vaccines An alternative approach to the development of live vaccines is to start with the food-poisoning organism Salmonella typhimurium. This organism can be attenuated by the introduction of lesions in the aro genes, which encode enzymes involved in the biosynthesis of aromatic amino acids, paminobenzoic acid and enterochelins. However, the mutant strains can establish selflimiting infections in the mice and can be detected in low numbers in organs such as the liver and spleen. Already a wide range of heterologous antigens have been expressed in such vaccine strains (Hackett 1993). An alternative vector is the human oral commensal Streptococcus gordonii (Fischetti et al. A different procedure for attenuating a bacterial pathogen to be used as a vaccine has been proposed by Kaper et al. A restriction-endonuclease fragment encoding the A1, but not the A2 or B, sequence was deleted in vitro from cloned cholera-toxin genes. The resulting strain produces the immunogenic but non-toxic B subunit of cholera toxin but is incapable of producing the A subunit. This strain has been found to be safe and immunogenic in carefully controlled clinical trials in a number of countries (Cryz 1992). They wanted to develop a vaccine against group B meningococci but were hampered by the fact that there was considerable sequence variation in surface-exposed proteins. Starting with the entire genome sequence of a group B strain of Neisseria meningitidis, they identified 350 proteins as potential protective antigens. The sera from the mice allowed the identification of proteins that are surfaceexposed in meningococci, that are conserved across a range of strains and that induce a bactericidal antibody response, a property known to correlate with vaccine efficacy in humans. Recombinant viruses as vaccines Recombinant viruses can be used as vectors to express heterologous antigens, and thus function as live vaccines. The first animal virus to be exploited in this way was vaccinia, which had been used previously as a non-recombinant vaccine providing cross-protection against variola virus, the causative agent of smallpox. Contributing to its success as a live vaccine were its stability as a freeze-dried preparation, its low production cost and the ability to administer the vaccine by simple dermal abrasion. Vaccinia remains the most widely explored recombinant viral vaccine, and many antigens have been expressed using this vector, essentially using the methods described on p. The most successful recombinant-virus vaccination campaign to date involved the use of recombinant vaccinia virus expressing rabies-virus glycoprotein. This was administered to the wild population of foxes in central Europe by providing a bait consisting of chicken heads spiked with the virus. The epidemiological effects of vaccination were most evident in eastern Switzerland, where two decades of rabies came to a sudden end after only three vaccination campaigns (Brochier et al. One disadvantage of this approach in humans is the unacceptably high risk of adverse reactions to the vaccine. For use in humans, vaccinia must be further attenuated to make it replication-deficient and to minimize the likelihood of replication-competent viruses arising by recombination. Other poxviruses, such as canarypox, have also been developed as potential vaccines. As well as vaccinia, adenovirus and alphavirus vectors have also been investigated as potential recombinant vaccines. Antigen Rabies virus glycoprotein Vesicular stomatitis glycoprotein Herpes simplex virus glycoprotein D Hepatitis B surface antigen Influenza virus haemagglutinin Human immunodeficiency virus envelope Reproduced with permission from Tartaglia & Paoletti (1988). Neutralizing antibodies + + + + + + Cellular immunity + + + + + + Animal protection + + + + + not determined Table 14. However, stronger stimulation of the immune response can often be achieved by presenting the antigen on the surface of the virus attached to a host virus-derived carrier protein. The advantage of this strategy is that the recombinant antigen is presented as multiple copies. This strategy is borrowed from the surface display of foreign antigens on bacteria that are used as live vaccines (see above) but is much safer. Indeed, while a number of conventional viral vaccines, such as vaccinia, have been developed as surface-display systems. Plants as edible vaccines Plants have been explored as a cheap, safe and efficient production system for subunit vaccines (Table 14. The earliest demonstration was the expression of a surface antigen from the bacterium S. This bacterium is the causative agent of dental caries, and it was envisaged that stimulation of a mucosal immune response would prevent the bacteria colonizing the teeth and therefore protect against tooth decay (Curtis & Cardineau, 1990). A number of edible transgenic plants have been generated expressing antigens derived from animal viruses. As well as animal virus antigens, autoantigens associated with diabetes have also been produced (Ma et al. Plants have also been infected with recombinant viruses expressing various antigen epitopes on their surfaces. The first clinical trials using a plant-derived vaccine were conducted in 1997 and involved the ingestion of transgenic potatoes expressing the B subunit of the E. This resulted in a successful elicitation of mucosal immunity in test subjects (Tacket et al. In each of the examples above, the functional component of the vaccine introduced into the host is the protein responsible for the elicitation of the immune response. The advantages of this method include its simplicity, its wide applicability and the ease with which large quantities of the vaccine can be produced. Selecting targets for new antimicrobial agents In attempting to develop new antimicrobial agents, including ones that are active against intractable pathogens such as the malarial parasite, it would be useful to know which genes are both essential for virulence and unique to the pathogen. Once these genes have been identified, chemical libraries can be screened for molecules that are active against the gene product. Secondly, target genes can be selected on the basis that there are no human counterparts. Grow in presence of thymine Isolate pathogen on media + thymine + Xgal White colonies thyA expressed from promoter only active in vivo Blue colonies thyA expressed from promoter active in vitro. The pool of fusions is transferred to a strain of the pathogen carrying a purA or thyA deletion and selection made for integration into the chromosome. Fusions that contain a promoter that is active in the animal allow transcription of the purA or thyA gene and hence bacterial survival. When the surviving pathogens are reisolated from the test animal, they are tested in vitro for their levels of -galactosidase. The tnpR locus encodes a site-specific resolvase and is used as the selection gene. Expression of this gene results in resolvase synthesis and the deletion of a resolvase-specific reporter. The advantage of this system is that it can be used to identify those genes that are expressed only transiently during animal infection (Camilli & Mekalanos 1995, Camilli 1996). Differential fluorescence induction this technique represents a different way of identifying environmentally controlled promoters and was originally developed to facilitate identification of Salmonella genes that are differentially expressed with macrophages (Valdivia & Falkow 1997). Bacteria that were non-fluorescent in the extracellular environment were sorted and used for a second round of macrophage infection. Signature-tagged mutagenesis this technique is a variation on the use of transposon mutagenesis that has been applied to the identification of bacterial virulence genes. Although one could test each mutant individually, this would be very laborious and would use large numbers of animals. By using signature-tagged mutagenesis, one can test large numbers of different mutants simultaneously in the same animal. The use of sequence-tagged mutagenesis was first described for the identification of virulence genes from S. The double-stranded tags were ligated into a Tn5 transposon and transferred from E. A library of over 1500 exconjugants resulting from transposition events was stored in microtitre dishes. Each pool was injected into the peritoneum of a different mouse and infection allowed to proceed. Bacteria were then recovered from each mouse by plating spleen homogenates on culture media. Essentially, these techniques are methods for scanning the entire microbial genome for genes that are expressed under particular environmental conditions. Theme 4: Protein engineering 4 Conjugate plasmids into Salmonella and select for transposition of tagged transposon. The simplest example of protein engineering involves site-directed mutagenesis to alter key residues, as originally shown by Winter and colleagues (Winter et al. Thus Perry and Wetzel (1984) were able to increase the thermostability of T4 lysozyme by the introduction of a disulphide bond. However, although new cysteine residues can be introduced at will, they will not necessarily lead to increased thermal stability (Wetzel et al. Improving therapeutic proteins with single amino acid changes As noted earlier (p. After period for infection to be established, re-isolate kanamycin resistant cells from mouse. Tags that are missing must be attached to transposons that have inserted into a gene essential for mouse virulence. In this way 28 different mutants with attenuated virulence were identified and some of these mutants were in genes not previously identified. The principle of signature-tagged mutagenesis has been extended to the analysis of pathogenicity determinants in a wide range of bacteria (for a review, see Handfield & Levesque 1999) and to fungi (Brookman & Denning 2000). For example, in the neutral solutions used for therapy, insulin is mostly assembled as zinc-containing hexamers. Not only have these insulins preserved their biological activity, but they are also absorbed two to three times faster. This in turn significantly increased the circulatory half-life, which in the native enzyme is only 5 min (Lau et al. Improving enzymes: subtilisin as a paradigm for protein engineering Proof of the power of gene manipulation coupled with the techniques of in vitro (random and sitedirected) mutagenesis as a means of generating improved enzymes is provided by the work done on subtilisin over the past 15 years (for review, see Bryan 2000). Every property of this serine protease has been altered, including its rate of catalysis, substrate specificity, pH-rate profile and stability to oxidative, thermal and alkaline inactivation. In the process, well over 50% of the 275 amino acids of subtilisin have been changed. At some positions in the molecule, the effects of replacing the usual amino acid with all the other 19 natural amino acids have been evaluated. Smokers are more prone to emphysema, because smoking results in an increased concentration of leucocytes in the lung and consequently increased exposure to neutrophil elastase.
These reclassifications and restatements have no impact on previously reported results of operations or retained earnings treatment for dogs dry eye discount myambutol 600 mg overnight delivery. The Companies use derivative transactions of antibiotics erectile dysfunction myambutol 800mg overnight delivery, for example antibiotic z pak discount 600 mg myambutol with amex, forward currency exchange and currency swaps that are used to hedge the risk of fluctuation in foreign currency exchange rates with respect to monetary receivables and payables denominated in foreign currencies resulting from import and export transactions bacteria estomacal generic 400mg myambutol fast delivery. With respect to other derivative transactions antimicrobial examples purchase 400mg myambutol overnight delivery, interest rate swap transactions are used to hedge the risk of fluctuation in interest rates antimicrobial resistance fda buy 800mg myambutol overnight delivery. The Companies evaluate hedge effectiveness by comparing the cumulative changes in cash flows from, or the changes in fair value of, hedged items with the corresponding changes in the hedging derivative instruments. The Companies report periodically to the Chief Financial Officer and the Treasury Office on the actual results of derivative transactions. Furthermore, the Companies enter into contracts with banks and securities houses with high credit rating to minimize credit risk exposure. The Companies principally enter into derivative transactions in connection with managing their market risk and not for speculation or trading purposes. To manage that risk, the Companies check the balance of the accounts and confirm the collection of money at the due date. Marketable securities are negotiable certificates of deposit subject to settlement in the short term. Securities are exposed to market price fluctuation risk; however, the Companies only hold shares in firms with which they have business relations and these are not held for speculation. Short-term loans receivable are used mainly for operating purposes, and funding through corporate bonds and long-term loans payable is mainly for capital investment. Debts with a floating rate are exposed to interest rate fluctuation risk, but (c) Supplementary information on fair values the fair value of financial instruments is calculated based on quoted market price or, in cases where there is no market price, by making a reasonable estimation. Because the preconditions applied include a floating element, estimations of fair value may vary. The contracted amounts, as presented in Note 6, "Derivative transactions," do not reflect market risk. Items for which fair value is difficult to estimate are not included in the following tables. Derivative transactions are presented net of receivables and liabilities, and figures within parenthesis indicate net liabilities. The terms of negotiable certificates of deposit are short term and the fair value thereof is nearly equal to book value, so the book value is used as fair value. See Note 5, "Investment securities" for information on investment securities categorized by holding purpose. Moreover, the fair value of long-term loans receivable that are doubtful is estimated in the same way or is provided in an amount sufficient to cover possible losses on collection. Liabilities (1) Payables and (2) short-term loans payable the terms of all of the above are short term and the fair value thereof is nearly equal to book value, so the book value is used as fair value. In cases where there is no market price, fair value is calculated by using the discounted cash flow based on the sum of the principal and total interest of the remaining period and credit risk. Certain long-term loans payable with floating rates are tied to interest rate swap transactions and subject to conventional treatment. Note 6 Derivative transactions (1) the following tables summarizes market value information of outstanding derivative transactions as of March 31, 2014 for which hedge accounting is not applied. Outstanding positions, for which gains and losses were recognized in the consolidated financial statements as of March 31, 2014, were as follows: Currency-related derivatives Millions of yen 2014 Contract amount Amount of principal due over one year Fair value Recognized gain (loss) Foreign currency swap transactions: Japanese yen received for Euro U. Currency-related derivatives: Principle-based method Millions of yens 2014 Contract amount Amount of principal due over one year Fair value Foreign currency forward contract transactions: Sell: U. The fair value of foreign currency swap transactions and interest rate swap transactions is based on the prices presented by the counterpart financial institutions. Outstanding positions, for which gains and losses were recognized in the consolidated financial statements as of March 31, 2015, were as follows: Currency-related derivatives Millions of yen 2015 Contract amount Amount of principal due over one year Fair value Recognized gain (loss) Foreign currency swap transactions: Japanese yen received for Euro U. Currency-related derivatives: Principle-based method Millions of yen 2015 Contract amount Amount of principal due over one year Fair value Foreign currency forward contract transactions: Sell: U. Therefore, the fair value of interest rate swaps is included in the fair value of the hedged long-term debt. Note 7 Inventories Inventories at March 31, 2014 and 2015 consisted of the following: Millions of yen Thousands of U. Long-term loans payable at March 31, 2014 and 2015 consisted of the following: Millions of yen Thousands of U. The funded status of the multiemployer contributory funded pension plans at December 31, 2014 (based on information available as of March 31, 2015), for which contributions are recorded as net periodic retirement benefit costs by the Companies, is as follows: Millions of yen Thousands of U. Note 10 Net assets earnings reserve is included in retained earnings in the accompanying consolidated balance sheets. All additional paid-in capital and all legal earnings reserve may be transferred to other capital surplus and retained earnings, respectively, which are potentially available for dividends. The maximum amount that the Company can distribute as dividends is calculated based on the unconsolidated financial statements of the Company in accordance with Japanese laws and regulations. Under Japanese laws and regulations, the entire amount of the issue price of shares is required to be accounted for as common stock, although a company may, by resolution of its Board of Directors, account for an amount not exceeding one-half of the issue price of the new shares as additional paid-in capital. Under the Japanese Corporate Law, in cases where dividend distribution of surplus is made, the smaller of an amount equal to 10% of the dividend and excess, if any, of 25% of common stock over the total of additional paid-in capital and legal earnings reserve must be set aside as additional paid-in capital or legal earnings reserve. Recoverable amount was measured by selling price, based on real estate appraisals or similar methods. If it is determined that an idle asset cannot be sold or diverted to another use, the asset is valued at zero. Certain consolidated subsidiaries accounted for impairment losses for the year ended March 31, 2015 as follows: Impairment loss Location Usage purpose Type of assets Millions of yen Thousands of U. Tokuyama City in Yamaguchi Prefecture Others Performance Polymer Products facilities Machinery, etc. Due to loss before income tax for the year ended March 31, 2015, reconciliation between the effective tax rate and the actual income tax rate is omitted. The Company is subject to a number of taxes based on income, which, in the aggregate, indicate a statutory rate in Japan of approximately 37. Accordingly, Special Corporate Tax for Reconstruction will no longer be imposed from the fiscal year commencing on or after April 1, 2014. Thus, the effective statutory tax rate used to calculate the deferred tax assets and deferred tax liabilities for the current fiscal year was changed from 37. Consequently, the amount of deferred tax assets (less the amount of deferred tax liabilities) declined by 352 million yen, income taxes-deferred posted in the current fiscal year increased by 356 million yen while deferred gains (losses) on hedges increased by 4 million yen. Accordingly, the income tax rate and other related matters for fiscal years commencing on or after April 1, 2015 have been revised. Thus, the effective statutory tax rate used to calculate deferred tax assets and deferred tax liabilities was changed from the previous 35. The effective statutory tax rate used to calculate deferred tax assets and deferred tax liabilities was changed from 35. Furthermore, the deduction limit for deductions of loss carryforwards has been amended to an amount equivalent to 65% of taxable income before deduction of loss carryforwards for fiscal years commencing on or after April 1, 2015, and an amount equivalent to 50% of taxable income before deduction of loss carryforwards for fiscal years commencing on or after April 1, 2017. Teijin Limited the account and the amounts related to stock options in the years ended March 31, 2014 and 2015 are as follows: Millions of yen Thousands of U. Non-exercisable stock options Stocks Company name Stock options outstanding at April 1, 2014 Stock options granted Forfeitures Conversion to exercisable stock options Stock options outstanding at March 31, 2015 Infocom Corporation 2014 2015 - - - - - - 23,000 - 23,000 - Exercisable stock options Stocks Company name Stock options outstanding at April 1, 2014 Conversion from non-exercisable stock options Stock options exercised Forfeitures Stock options outstanding at March 31, 2014 the following table summarizes value information of stock options as of March 31, 2015. Up to and including the year ended March 31, 2015, the Company has divided its operations into business groups based on the type of product, nature of business and services provided. The business groups formulate product and service strategies in a comprehensive manner in Japan and overseas. Accordingly, the Company divided its operations into four reportable operating segments on the same basis as applied internally: Advanced Fibers and Composites; Electronics Materials and Performance Polymer Products; Healthcare; and Trading and Retail. The description of each segment is as follows: Advanced Fibers and Composites: - Production and sales of aramid fibers, carbon fibers, polyester fibers and composites for industrial applications Electronics Materials and Performance Polymer Products: - Production and sales of films and resins for various industrial applications Healthcare: - Production and sales of prescription and non-prescription drugs and production, sales and rental of home healthcare devices Trading and Retail: - Trading and retail of polyester filaments, other fibers and polymer products (2) Accounting methods used to calculate segment sales, segment income, segment assets and other items for reportable operating segments Accounts for reportable operating segments are for the most part calculated in line with generally accepted standards for the preparation of consolidated financial statements. Amounts for intersegment transactions or transfers are calculated based on market prices or on prices determined using the cost-plus method. As described in Note 2, the Company recognizes certain of the home healthcare devices as fixed assets and depreciates them using the straight-line method. Reconciliations of published figures and aggregates of reportable operating segments as of March 31, 2014 and 2015 are as shown below: Millions of yen Thousands of U. Where noted, other italicized product names and service names used in this report are protected as the trademarks and/or trade names of other companies. Clearly describing the important metabolic processes that occur in prokaryotes under different conditions and in different environments, this advanced text provides an overview of the key cellular processes that determine bacterial roles in the environment, biotechnology and human health. Prokaryotic structure and composition are described as well as the means by which nutrients are transported into cells across membranes. The regulation of metabolism through control of gene expression and enzyme activity is also covered, as well as the survival mechanisms used under starvation conditions. Professor Byung Hong Kim is an expert on anaerobic metabolism, organic degradation and bioelectrochemistry. He has carried out research at several universities around the world, with an established career in the Korea Institute of Science and Technology. He has been honoured by the Korean Government, which designated his research group a National Research Laboratory, the Bioelectricity Laboratory, and has served as President of the Korean Society for Microbiology and Biotechnology. Professor Kim wrote the classic Korean microbiology text on Microbial Physiology and has published over 100 refereed papers and reviews, and holds over 20 patents relating to applications of his research in environmental biotechnology. Professor Geoffrey Michael Gadd is an authority on microbial interactions with metals, minerals and radionuclides and their applications in environmental biotechnology. He holds a personal Chair in Microbiology at the University of Dundee and is the Head of the Division of Molecular and Environmental Microbiology in the College of Life Sciences. He has published over 200 refereed scientific papers, books and reviews and has received invitations to speak at international conferences in over 20 countries. Professor Gadd has served as President of the British Mycological Society and is an elected Fellow of the Institute of Biology, the American Academy of Microbiology, the Linnean Society, and the Royal Society of Edinburgh. He has received the Berkeley Award from the British Mycological Society and the Charles Thom award from the Society for Industrial Microbiology for his outstanding research contributions to the microbiological sciences. Subject to statutory exception and to the provision of relevant collective licensing agreements, no reproduction of any part may take place without the written permission of Cambridge University Press. Prokaryotic organisms include bacteria and archaea and, although remaining relatively small and simple in structure throughout their evolutionary history, exhibit incredible diversity regarding their metabolism and physiology. Such metabolic diversity is reflective of the wide range of habitats where prokaryotes can thrive and in many cases dominate the biota, and is a distinguishing contrast with eukaryotes that exhibit a more restricted metabolic versatility. Thus, prokaryotes can be found almost everywhere under a wide range of physical and chemical conditions, including aerobic to anaerobic, light and dark, low to high pressure, low to high salt concentrations, extremes of acidity and alkalinity, and extremes of nutrient availability. The explosion of knowledge resulting from the development and application of molecular biology to microbial systems has perhaps led to a reduced emphasis on their physiology and biochemistry, yet paradoxically has enabled further detailed analysis and understanding of metabolic processes. Almost in a reflection of the bacterial growth pattern, the number of scientific papers has grown at an exponential rate, while the number of prokaryotic genome sequences determined is also increasing rapidly. This production of genome sequences for a wide range of organisms has made an in-depth knowledge of prokaryotic metabolic function even more essential in order to give biochemical, physiological and ecological meaning to the genomic information. Our objective in writing this new textbook was to provide a thorough survey of the prokaryotic metabolic diversity that occurs under different conditions and in different environments, emphasizing the key biochemical mechanisms involved. We believe that this approach provides a useful overview of the key cellular processes that determine bacterial and archaeal roles in the environment, biotechnology and human health. We concentrate on bacteria and archaea but, where appropriate, also provide comparisons with eukaryotic organisms. It should be noted that many important metabolic pathways found in prokaryotes also occur in eukaryotes further emphasizing prokaryotic importance as research models in providing knowledge of relevance to eukaryotic processes. In the second part, the trophic variations found in prokaryotes are described, including the use of organic compounds other than glucose, anaerobic fermentation, anaerobic respiration, chemolithotrophy and photosynthesis. In the third part, the regulation of metabolism through control of gene expression and enzyme activity is covered, as well as the survival mechanisms used by prokaryotes under starvation conditions. This text is relevant to advanced undergraduate and postgraduate courses, as well as being of use to teachers and researchers in microbiology, molecular biology, biotechnology, biochemistry and related disciplines.
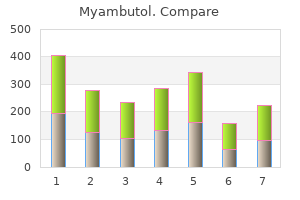
Order myambutol 600 mg amex. HSN | Copper Fit 12.30.2016 - 08 PM.
The most common form of O-glycan attached to glycoproteins from mammalian cells is the mucin-type antimicrobial activity purchase myambutol 400 mg with amex, which involves the addition of N-acetylgalactosamine to a serine or threonine residue in a protein how does antibiotics for acne work myambutol 400mg free shipping. No consensus sequence has been identified antibiotic resistance methods purchase myambutol 600mg free shipping, although glycosylation often occurs in a region of the protein that contains a high proportion of serine antibiotics haven't worked for uti cheap myambutol 400 mg, threonine antibiotics for sinus infection purchase myambutol 400 mg online, and proline (Van den Steen et al infection 7 days to die generic myambutol 800mg. Further elongation leads to a large number of structures, synthesized by various glycosyltransferases, producing eight different core structures (Figure 6. These core structures can be further modified by sialylation, fucosylation, sulfation, methylation, or acetylation. These cells can be manipulated genetically and show high productivity in large-scale systems. However, the cells lack any metabolic pathways for glycosylation and so the proteins produced are not glycosylated. Lower eukaryotes (yeast, insect, and plant cells) are capable of glycosylation of proteins. However, the glycans produced in these cells differ significantly from those present in mammalian glycoproteins (Jenkins et al. These represent a series of characteristic structures attached to a serine (S) or threonine (T) amino acid residue of a protein. There are eight core structures, each of which may have further monosaccharide additions. Although there is extensive heterogeneity of structures arising from any cell type, examples of predominant N-glycans that might occur from different systems are shown in Table 6. Lepidopteran insect cell lines (such as Spodoptera frugiperda, Sf-9) have been used extensively for expression of recombinant proteins using the baculovirus as an expression vector (Jarvis et al. Alternative methods of protein expression are also available using efficient insect-associated promoter systems (Farrell et al. The advantage of the use of these cells is the high expression level and growth rate of the cells in culture. Glycosyltransferase enzymes are either absent or at a low level of activity (Jarvis et al. Therefore, generally the insect expression system is incapable of synthesizing sialylated lactosamine complex-type N-glycan or sialylated O-glycans. N-glycan None High mannose Structure - Insect Fucosylated core structure Plant Xylosylated and fucosylated core structure Complex biantennary Mammalian 136 Animal Cell Technology Jarvis et al. Depending upon the relative activities of these competing processing enzymes, trimannosyl core or complex N-linked glycans may be produced. The enzymic activity may vary considerably and this explains the differences in the N-glycan pattern of the same glycoprotein secreted by different insect cell lines (Kulakosky et al. The potential of insect cells for O-glycosylation was reported in a study in which three lepidopteran cell lines were shown to produce predominantly short O-glycan structures (Lopez et al. There was no indication of sialylation of these structures, suggesting the absence of sialyltransferase activity. The culture medium used to grow these cell lines had a major effect on the O-glycans expressed. The use of a semi-defined rich medium enhanced glycosyltransferase activity significantly compared with a minimal nutrient medium. The limitations in the ability of insect cells for glycosylation have restricted their use for production of human therapeutics. Genetic engineering of these systems is under study in an attempt to improve the glycosylation machinery to produce more ``humanized' glycoproteins (Jarvis et al. The early steps in the addition of carbohydrate to proteins appear to have been remarkably conserved during evolution. However, although relatively few N-linked sites have been characterized, it was noted that there is a trend in favor of the use of Asn-X-Thr sites over Asn-X-Ser in yeast glycoproteins. Most yeast and filamentous fungi synthesize carbohydrate chains of the high mannose type. Addition of Man residues to core oligosaccharides occurs very rapidly in the Golgi, forming the characteristic high mannose structures (mannan), which can consist of more than 50 mannose residues and resulting in high molecular weight glycoproteins (Hersecovics and Orlean, 1993). Post-translational modification of recombinant proteins 137 Proteins synthesized in yeast may also contain O-glycans consisting of linear poly-mannose structures attached to Ser or Thr. Similar to mammalian cells, O-glycosylation in yeasts has no obvious consensus sequence. However, unlike mammalian cells O-glycosylation in yeast is initiated with covalent attachment of mannose via a dolichol phosphate mannose precursor. However, the processing of the oligosaccharide trimming and further modification of glycans in the Golgi differ from mammalian cells. Plantderived oligosaccharides do not possess sialic acid and frequently contain xylose (Xyl), not normally present in mammalian N-linked oligosaccharides. Differences in the relative activity of these enzymes among species can account for significant variations in structure. The fact that glycoproteins normally exist as mixtures of glycoforms suggests that the protein structure is not the primary determining factor in glycosylation. The glycoforms that emerge from the Golgi are end products of a series of incomplete enzymic reactions. Thus, the choice of the host cell line is a particularly important factor in the glycoform profile of a recombinant protein (Rudd and Dwek, 1997). Sialylation patterns of the secreted protein are particularly affected by the host cell. However, this may not be so easy given that the extent of glycosylation may decrease over time in a batch culture (Curling et al. This is likely to be due to the continuous depletion of nutrients (particularly glucose or glutamine) and accumulation of metabolic byproducts, which have been shown 138 Animal Cell Technology to limit the glycosylation process (Hayter et al. Culture conditions such as nutrient content, pH, temperature, oxygen, or ammonia, may have a significant impact on the distribution of glycan structures found on the resulting recombinant protein (microheterogeneity). This of course is of major concern in trying to produce consistent biopharmaceuticals. It can lead to enhanced glycoform heterogeneity and significant batch to batch variation in the production process. To maintain product quality it is important to understand the parameters that cause the variation in glycosylation. Glucose starvation may result in an intracellular depleted state with a shortage of glucose-derived precursors of glycans that could result in reduced site occupancy. Fed-batch strategies may be designed to insure that the concentrations of these key nutrients do not decrease below a critical level that could compromise protein glycosylation (Xie and Wang, 1997). Nutrient levels below these critical concentrations led to decreased sialylation and an increase in hybrid and high mannose-type glycans. This may be the mechanism for the observed decrease in sialylation that has been shown to accompany high levels of ammonia in culture (Yang and Butler, 2000). Terminal galactosylation of an immunoglobulin (IgG) was changed significantly with a gradual decrease in the digalactosylated glycans (G2) from 30% at the higher oxygen level to around 12% under low oxygen conditions (Kunkel et al. This introduces an extra negative charge in the protein that can result in alterations of properties. Low levels of deamidation can lead to protein aggregation and are related to amyloid formation (Nilsson et al. Deamidation may occur during bioprocessing or storage of biopharmaceuticals and is dependent upon a number of factors including pH, temperature, solvent dielectric constant, and ionic strength, as well as the structure of the protein. This may lead to loss of function or structural alterations of the proteins that are associated with the loss of a positive charge. Glutamic semialdehyde (5 mmole/mol) accounted for the majority of carbonyl groups in proteins from HeLa cells in culture. It is a complex process that results in lysine- and arginine-derived glycation adducts in intracellular or extracellular proteins. Biopharmaceuticals may be subjected to heat treatment as a means of viral inactivation. Protein damage is prevented by the addition of high concentrations of a thermostabilizing excipient such as sugars or polyhydric alcohols. The non-reducing polyhydric additives increase thermostability of the protein by the formation of a hydrogen-bonded solvent shell. Glucose or other sugars may react non-enzymatically with proteins to form glycation products. Such a treatment will destroy most viruses but may cause glycation products that could render the protein immunogenic (Smales et al. The observed modifications take place by reaction adducts of glucose or fructose that are derived from the hydrolysis of sucrose. The extra negative charge provided by the carboxylation aids in the calcium-induced interaction of these proteins with membrane surfaces. However, a higher secretion rate decreased the efficiency of gamma-carboxylation (Guarna et al. Similarly, culture conditions were shown to affect carboxylation of protein C, with a good supply of oxygen favoring high efficiency of gamma-carboxylation (Sugiura and Maruyama, 1992). Factor X undergoes extensive post-translational modification including gamma-carboxylation of 11 glutamic acid residues. However, the efficiency of this process is variable and inevitably leads to a heterogeneity of isoforms. This may depend on the availability and activity of the carboxylase enzyme that binds to the propeptide. Surprisingly, this increased the extent of carboxylation from a normal value of 32% to 85% by enhancing substrate turnover. Nevertheless, in all situations there were two pools of protein produced: a fully carboxylated and uncarboxylated recombinant factor X. This is an enzymic reaction catalyzed by a carboxylase to modify a glutamic acid residue of a protein to form gamma-carboxyglutamic acid. For mAbs the C-terminal lysine of heavy chains may be removed, giving rise to a mixture of Lys0 (absence) or Lys1 (presence) heavy chain variants. It is considered that the removal of either C-terminal lysine or arginine is unlikely to cause loss of biological activity because they are found as natural variants of these proteins under physiological conditions in vivo. This enzyme can convert proline to hydroxyproline when the substrate is contained in an amino acid motif (X-Pro-Gly). This is known to occur in collagens and requires four co-substrates: ferrous ion, 2-oxoglutarate, oxygen, and ascorbate (Kivirikko et al. An example of this type of conversion has also been found in the N-terminus of the prion protein in which there is a domain containing poly (L-proline) (Gill et al. The post-translational modification of these proteins is essential to insure structural stability and biological and clinical activity. This gives rise to a significant heterogeneity of structural forms and it is important to monitor these to insure consistency of production. However, the ability to control the glycosylation is limited by our understanding of the parameters that affect the heterogeneity of added glycan structures. It is clear that the glycosylation process is affected by a number of factors including the three-dimensional structure of the protein, the enzyme repertoire of the host cell, the transit time in the Golgi, and the availability of intracellular sugar-nucleotide donors. From a process development perspective there are many culture parameters that can be controlled to enable a consistent glycosylation profile to emerge from each batch culture. A further, but more difficult goal is to control the culture Post-translational modification of recombinant proteins 143 conditions to enable the enrichment of specific glycoforms identified with desirable biological activities. There are also a number of other possible post-translational modifications of proteins that can be characterized as the addition or removal of small organic residues. These may be important for the structural integrity of the protein and should be monitored in culture bioprocesses designed for biopharmaceutical production. References Akagawa M, Sasaki T, Suyama K (2002), Oxidative deamination of lysine residue in plasma protein of diabetic rats. Differences in glycosylation and proteolytic processing lead to heterogeneity in batch culture, Biochem. Gavel Y, von Heijne G (1990), Sequence differences between glycosylated and nonglycosylated Asn-X-Thr/Ser acceptor sites: implications for protein engineering, Protein Eng. Kornfeld R, Kornfeld S (1985), Assembly of asparagine-linked oligosaccharides, Annu. Lopez M, Tetaert D, Juliant S, Gazon M, Cerutte M, Verbert A, Delannoy P (1999), O-glycosylation potential of lepidopteran insect cell lines, Biochim. Maras M, Saelens X, Laroy W, Piens K, Claeyssens M, Fiers W, Contreras R (1997), In vitro conversion of the carbohydrate moiety of fungal glycoproteins to mammalian-type oligosaccharides. Evidence for N-acetylglucosaminyltransferase-Iaccepting glycans from Trichoderma reesei.
These reactions are (1) transfer of energy to the adjacent molecule exciting its electron (resonance transfer) antibiotic treatment for sinus infection purchase myambutol 400mg fast delivery, (2) oxidation of the excited pigment transferring an electron to a second compound or (3) emission of fluorescent light bacterial vaginosis buy generic myambutol 400 mg on-line. When a photon is radiated to a surface antibiotic resistance methods buy myambutol 600mg with mastercard, it may be reflected antibiotic of choice for uti order myambutol 600mg with visa, transmitted bacteria use restriction enzymes to quizlet purchase 600mg myambutol with visa, or absorbed kinds of antibiotics for acne myambutol 600 mg without a prescription. Different pigments absorb photons of different wavelengths depending on the nature of the absorbing pigment. When a photon is absorbed by a pigment, the energy of light is converted to kinetic energy in the form of an excited electron. When a molecule (or atom) absorbs a photon an electron is boosted to a higher energy level by transferring an electron from the normal shell to the outer shell (Figure 11. These reactions are (1) transfer of energy to the adjacent molecule exciting its electron (resonance transfer), (2) reduction of a second compound (oxidation, photo-induced charge separation) or (3) emission of fluorescent light. The energy gained by the pigments through such excitation is referred to as the exciton. The exciton is transferred from antenna molecules to the reaction centre through resonance transfer, and this reaction takes about 0. The electron transport reaction at the reaction centre is initiated by the oxidation with the consumption of resonance energy. Resonance transfer takes place from molecules of a higher exciton to those with a lower exciton. The oxidized chlorophylls are reduced again, either oxidizing externally supplied electron donor(s) in a process known as non-cyclic electron transport or by taking the original electrons through the cyclic electron transport system. Chlorophyll a serves as the photosynthetic pigment in the reaction centres of both photosystems. The differences in the maximum absorption of chlorophyll a are due to the proteins forming the complex. Electrons are transferred from A0 to ferredoxin through phylloquinone and several [Fe-S] centres. The electrons from the reduced ferredoxin are transferred back to P700 through the cytochrome bf complex and plastocyanin in a process known as cyclic electron transport (Figure 11. Electrons are transferred from A0 to ferredoxin through phylloquinone (A1) and several [Fe-S] centres. The free energy changes in cyclic electron transport are conserved as a proton motive force transporting protons into the thylakoid. Green bacteria have only cyclic electron transport, and reduced sulfur or organic compounds serve as the electron donor. On the other hand, the other members of the green bacteria, the filamentous anoxygenic phototrophic bacteria, can grow chemoorganotrophically under aerobic conditions. The latter have a different photosynthetic electron transport system from that of the green sulfur bacteria which is similar to that of the purple bacteria (see Figure 11. Heliobacteria have P788 as the reaction centre, which is different from the green sulfur bacteria, although the photosynthetic mechanisms are similar. The excited P870 reduces bacteriopheophytin to begin the cyclic electron transport for the generation of a proton motive force. Sulfur and organic compounds are used as electron donors reducing cytochrome c2 (Figure 11. Purple sulfur bacteria cannot grow under aerobic conditions, and purple non-sulfur bacteria grow chemoorganotrophically under aerobic conditions. They have a similar cyclic electron transport system as the purple bacteria, as depicted in Figure 11. Under dark conditions, purple non-sulfur bacteria can grow with or without molecular oxygen. Heliobacteria and aerobic anoxygenic photosynthetic bacteria supplement their energy requirements through light reactions while growing as chemoorganotrophs. On the other hand, filamentous anoxygenic phototrophic bacteria can use organic electron donors in phototrophic metabolism and can grow chemoorganotrophically like purple non-sulfur bacteria. Obligately photolithotrophic cyanobacteria cannot use glucose, though they metabolize glycogen as storage materials with glycolytic enzymes. Figure 11:15 the structure of retinal bound to rhodopsins in species of Halobacterium. A group of proteins known as rhodopsins facilitate motility and photophosphorylation. Sensory rhodopsins with a similar structure pass the information to the transducer proteins to control phototaxis. At low dissolved O2 concentrations, the organisms move towards the light through the actions of sensory rhodopsin I and transducer I to use it. Bacteriorhodopsin and halorhodopsin convert light energy into a proton motive force. Evolutionary relationships among photosynthetic prokaryotes (Heliobacterium chlorum, Chloroflexus aurantiacus, cyanobacteria, Chlorobium tepidum and proteobacteria): implications regarding the origin of photosynthesis. Suitability of different photosynthetic organisms for an extraterrestrial biological life support system. Adaptation and acclimation of photosynthetic microorganisms to permanently cold environments. Heterocyst differentiation and pattern formation in cyanobacteria: a chorus of signals. PpsR: a multifaceted regulator of photosynthesis gene expression in purple bacteria. Seeing green bacteria in a new light: genomics-enabled studies of the photosynthetic apparatus in green sulfur bacteria and filamentous anoxygenic phototrophic bacteria. Biliproteins and phycobilisomes from cyanobacteria and red algae at the extremes of habitat. Adaptive mechanisms of nitrogen and carbon assimilatory pathways in the marine cyanobacteria Prochlorococcus. Structure, function, regulation, and assembly of D-ribulose-1,5-bisphosphate carboxylase/oxygenase. Bacteriorhodopsin: a high-resolution structural view of vectorial proton transport. Organic materials are converted to carbon skeletons for monomer and polymer synthesis, as well as being used to supply energy. This is possible through the regulation of the reactions of anabolism and catabolism. With a few exceptions, microbial ecosystems are oligotrophic with a limited availability of nutrients, the raw materials used for biosynthesis. Furthermore, nutrients are not usually found in balanced concentrations while the organisms have to compete with each other for available nutrients. Unlike animals and plants, unicellular microbial cells are more directly coupled to their environment, which changes continuously. Many of these changes are stressful so organisms have evolved to cope with this situation. Since almost all biological reactions are catalyzed by enzymes, metabolism is regulated by controlling the synthesis of enzymes and their activity (Table 12. In this chapter, different mechanisms of metabolic regulation are discussed in terms of enzyme synthesis through transcription and translation and enzyme activity modulation. Various mechanisms regulating the synthesis of individual enzymes are discussed here before multigene regulation is considered. The group 1 D is essential, and responsible for the transcription of most genes in exponentially growing cells. A group 2 -factor, S, participates in the expression of stationary phase proteins. Group 3 -factors include F, responsible for flagella synthesis, and H, the heat shock response -factor in Escherichia coli. This group includes the extracytoplasmic function -factor subfamily, the largest and most divergent group. Genes for proteins needed in large quantities have strong promoters with a high affinity for the -factors. While the essential housekeeping D functions in exponentially growing cells, other -factors are activated according to the growth conditions, including E, F, H, N and S. The housekeeping -factor is called D in Gram-negative bacteria and A in Gram-positive bacteria. Consequently, -factors with the same name have different functions in different bacteria: D is the housekeeping -factor in Escherichia coli but is the -factor for flagella formation in Bacillus subtilis. In a related terminological problem, the extracytoplasmic function -factor, E, should not be regarded as functioning extracytoplasmically. Instead, E participates in the expression of genes needed to repair denatured proteins of extracytoplasmic location. Since a -factor recognizes multiple promoters throughout the chromosome, a specific -factor participates in the transcription of functionally unrelated genes. Proteins known as anti-sigma (anti-) factors bind -factors and inhibit their activity. When Escherichia coli is infected with bacteriophage T4, a phage-originating anti- factor, AsiA, inhibits the activity of the bacterial housekeeping D to produce more phage proteins. When E is required, either an anti-anti- factor binds RseA releasing E or RseA is degraded to release E. In the case of F responsible for flagellin synthesis, the anti- factor, FlgM, is exported into the environment through the flagellin export mechanism when more flagellin is needed (Section 12. Enzymes synthesized in the presence of substrates are referred to as inducible enzymes and the substrate is termed the inducer. Constitutive enzymes are those enzymes that are produced under all growth conditions. Inducible enzymes are generally those used in the catabolism of carbohydrates such as polysaccharides (cellulose, starch, etc. When a single inducer induces more than two enzymes, they are produced either simultaneously or sequentially. The former is referred to as coordinate induction, and the latter as sequential induction (Figure 12. Genes of coordinate induction are in the same operon, and genes from separate operons are induced sequentially. Figure 12:2 Coordinate induction and sequential induction of multiple enzymes by a single inducer. Springer, New York) Enzymes from genes of the same operon are induced simultaneously in coordinate induction (a) and genes from different operons are induced sequentially (b). The product of the first enzyme reaction is the inducer of the second enzyme in sequential induction. Their structural genes form an operon (lac operon) with a promoter and operator (Figure 12. The regulatory gene (lacI) next to the 50 end of the operon is expressed constitutively with its own promotor. On the other hand, when the inducer is available, it binds the repressor protein, removing it from the operator Figure 12:3 Induction mechanism of the lac operon in Escherichia coli. Springer, New York) the lac operon consists of a promoter, operator and structural genes, and a repressor protein is produced from the lacI gene that has its own promoter (a). In the absence of inducer, the repressor protein binds the operator region of the operons, preventing transcription of the structural genes (b1). The complex cannot bind the operator region, and the structural genes are transcribed (b2). Springer, New York) the regulator protein AraC is in the protein P1 form to bind the operator region, inhibiting transcription of the structural genes in the absence of arabinose (a). This mutant transcribes the structural genes as constitutive enzymes in the absence of the inducer. In this sense, the regulation by repressor proteins is referred to as negative control. Activator proteins are involved in the regulation of catabolic genes for arabinose, rhamnose, maltose and others. Genes for arabinose catabolism consist of araA, B, C, D, E, and F; araC is a regulatory gene encoding an activator protein. The term regulon is used to define genes of the same metabolism controlled by the same effectors scattered around the chromosome, such as ara genes. Regulation by an activator, as in the ara regulon, is referred to as positive control. This is due to the fact that the readily utilizable glucose and its metabolites repress the utilization of lactose. When Escherichia coli is cultivated on a mixture of glucose and lactose, the bacterium grows on glucose, repressing lactose utilization (A) at the beginning. When glucose is exhausted the bacterium grows again on lactose (C) after a lag period (B).